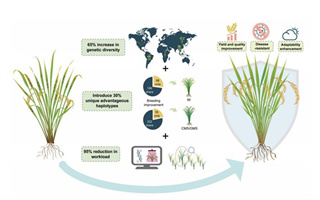
杂交水稻的商业化应用,中国从1970年代初就走到了世界的最前头,一直领跑至今,已经将近50年。半个世纪以来,突破了“三系法”,原创了“两系法”,在籼稻、粳稻亚种中,品种间杂种优势都得到了充分利用,造福了中国乃至世界部分人民。但是,品种间杂种优势这条“道”已经走了50年,育种家预定的主要目标(比如丰产、优质、抗病、耐热、抗倒等等)已经一个一个地实现,当前这条“道”已显得十分“拥挤”,前方的目标也越来越难实现,似乎有必要寻求新的“赛道”,以攀登新的高峰。本文从中国杂交水稻育种50年的实践和基础来推演和预判杂交水稻育种的“新赛道”,与业内人士交流。
PART.1
杂交水稻开创性研究的回顾
★1.1 国际早期研究
在国际上,美国农学家琼斯于1926年首先提出了水稻具有杂种优势的理论。1954年Sampath等报道水稻细胞质雄性不育现象。1958年日本东北大学Katsuo和Mizushima在粳稻亚种中开创水稻雄性不育系选育研究,用中国红芒野生稻与日本粳稻藤板5号杂交,经过连续回交,育成了世界第一个具有野生稻细胞质的粳稻藤板5号不育系。1966年日本琉球大学新城长友(Shinjyo)以印度籼稻Chinsuran Boro II为母本与台湾粳稻品种台中5号杂交再回交,1968年育成具有Chinsuran Boro II细胞质的台中5号粳稻不育系,也称包台型或BT型不育系,并实现粳稻“三系”配套。1968年日本农业技术研究所渡边用缅甸籼稻里德稻与日本粳稻藤板5号杂交,育成具有里德稻细胞质的粳稻藤板5号不育系。美国加州大学Erickson和Carnahan也分别于1969年和1972年发现台湾品种Brico和非洲光身稻都有导致加州粳稻雄性不育的细胞质。1970年韩国学者也育成具有野生稻细胞质的藤板5号不育系。1972年国际水稻研究所Vimani育成具有台湾本地1号细胞质的Pankhari 203不育系。
这些研究证明一点,水稻细胞质雄性不育性是一种普遍现象。但是,上述不育系不是由于找不到恢复系不能实现“三系”配套,就是虽然实现三系配套但优势不强或制种困难,均未能应用于商业化生产。
★1.2 中国的研究和突破
杂交水稻是中国对世界现代农业科技贡献的重大成就之一。袁隆平于1961年从天然杂交水稻获得灵感:自然界存在天然雄性不育株,从而开始寻找水稻天然雄性不育株。终于在1964年从洞庭早籼、南特号、胜利籼等籼稻品种中发现概率仅为1/50000的无花粉、花药退化或者花粉败育型的自然雄性不育株。然后,由全国多家水稻育种单位协作,通过大范围测交保持系和人工培育保持系等多种手段,历时6年与千百个水稻品种测交并筛选其雄性不育保持系,结果均达不到100%的保持不育,最终未能成功。后来研究证明,这些水稻雄性不育株均为普通雄性核不育株,难以找到保持系。
面对失败,袁隆平团队仍坚信当初的籼稻“三系”配套方案一定能实现,并改变策略,从亲缘关系较远的野生稻中寻找或通过远缘杂交创造雄性不育材料。直到1970年11月23日,袁隆平的助手李必湖在海南崖县南红农场沼泽地的普通野生稻群中发现一株花药瘦小、黄色、不开裂,内含典型花粉败育型雄性不育株,之后定名为野败。
1971年春多省研究人员从袁隆平团队获得野败不育株异交的种子或稻蔸,并对野败进行大量的测交。颜龙安团队由于意识到了野生稻的感光性,采用了遮光处理,野败不育株在江西正常抽穗开花,顺利开展了测交,找到保持品种二九矮1号、珍汕97等。袁隆平团队在1971年底也发现广矮3784等长江流域早籼种质能保持野败雄性不育。他们于1972年成功培育出二九南1号A、珍汕97A等系列野败型雄性不育系及其保持系二九南1号B、珍汕97B,至此才突破雄性不育保持系培育的难关。因此,科学界认为野败的发现为中国杂交水稻研究打开了突破口。
1972年朱英国团队在海南红芒野生稻同江西地方籼稻品种莲塘早杂交后代中发现雄性不育株,这是一种有别于野败型孢子体不育(典型败育花粉,花粉败育发生在单核后期)的新类型。雄性败育类型属于配子体不育(圆败型花粉,花粉败育发生在二核期)。后来,用莲塘早连续回交,于1974年成功培育出红莲型细胞质雄性不育系红莲A及其保持系。
1972年中国科学家引进包台型雄性不育资源,该品种是中国三系杂交粳稻应用最广的细胞质雄性不育资源。测交发现,绝大多数粳稻品种对该品种具有保持能力。第一个粳稻包台型(BT型)细胞质雄性不育系黎明A于1975年育成。
有了不育系和保持系,接下来就是找到强优势恢复系。从1972年冬天开始,“三系”育种主要集中在恢复系选育上。全国10多个省份的研究人员利用各类稻种资源进行了大量的测交恢复系试验。广西学者张先程、李丁民等发现野败细胞质雄性不育的恢复基因主要分布在血缘较近的低纬度热带稻种资源中,并从国际水稻研究所的IR系列品种中找到对野败雄性不育恢复较好的泰引1号、IR24、IR661、古154等种质;袁隆平于1973年10月宣告野败型雄性不育系、保持系和恢复系“三系”配套,次年育成南优2号、汕优2号等第一批野败型杂交水稻组合,打开了杂交水稻商业化应用的大门。
1973年湖北学者石明松在一季晚粳品种农垦58中发现3株自然雄性不育株,经过10多年的研究育成了光敏不育系农垦58S,建立了在长日照高温条件下进行杂交制种、短日照低温条件下自交繁殖不育系的光敏型“两系”法杂种优势利用技术体系。农垦58S不育性遗传行为复杂,控制其雄性育性的光敏核不育基因有pms1、pms2和pms3。其中pms3是位于第12号染色体上的一个非编码RNA基因的单碱基突变,该基因使长日照特异雄性育性关联RNA在长日照条件下转录量降低,并造成农垦58S花药程序化死亡提前,导致雄性不育。pms3是粳型光温敏核不育系的最重要不育基因源。1991年研究人员将农垦58S光敏核不育基因转育到籼稻中,育成第一个籼型光温敏核不育系培矮64S。1994年研究人员又育成第一个两系法杂交中籼组合。
1987年邓华凤等从籼稻三交组合超40B/H285//6209-3的F5群体中发现了1株在高温条件下表现雄性不育、低温条件下表现雄性可育的天然雄性不育株,继而育成籼型温敏不育系安农S-1,利用其温敏特性建立了在高温条件下进行杂交制种、低温条件下自交繁殖不育系的温敏型两系法杂种优势利用技术体系。安农S-1温敏核不育受第2号染色体上的隐性单基因tms5的调控。tms5编码一个RNA酶ZS1,它的第71位C→A的突变导致高温核雄性不育。该基因遗传行为简单,是目前中国两系法杂交水稻应用面积最大的遗传资源,已占两系法杂交水稻的80%以上。
野败的发现和红莲的育成,突破了三系法杂交水稻;农垦58S、安农S-1的发现和育成,原创了两系法杂交水稻,这些成果对杂交水稻的发展作出了奠基性的贡献。
PART.2
中国杂交水稻育种的成就和现状
★2.1 不育系的选育
不育系的选育是杂交水稻育种的第一关。自1972年颜龙安团队率先育成二九矮A、珍汕97A,次年袁隆平团队育出二九南1号A,至2021年12月21日,国家水稻数据中心收录通过省级以上审定的不育系572个,其中籼型三系不育系298个,籼型两系不育系227个;粳型三系不育系39个,粳型两系不育系8个。
从数量上可以看出,不育系的选育集中在籼稻,粳稻中选出的不育系较少,粳稻不育系数量不足籼稻不育系的10%。这或许与3个因素有关,一是我国粳稻的种植面积和育种力量远远小于籼稻;二是粳稻品种间杂种优势也不如籼稻品种间杂种优势大;三是粳稻常规品种的产量明显高于籼稻。所以,粳稻不育系选育和杂种优势利用远不如籼稻。同时,从数量上还可以看出,无论是籼稻还是粳稻,三系不育系的选育多于两系不育系。这或许是由于三系育种开始的时间长且育性更为稳定。
★2.2 不育系的分类及遗传学研究
关于植物雄性不育性理论,Sears最早于1947年提出“三型学说”,即细胞质基因控制的雄性不育、细胞质基因与细胞核基因互作控制的雄性不育、细胞核基因控制的雄性不育。按此理论,细胞质不育只有保持系,没有恢复系,因而没有利用价值;质核互作不育既有保持系,又有恢复系,可以用来生产杂交种;核不育没有保持系,只有恢复系,实际利用也没有可能。
后来的研究认为,纯粹由细胞质基因控制的雄性不育性可能并不存在,因此“三型学说”被Edwardson于1956年修改为“二型学说”,并将质核互作不育称之为细胞质不育,于是雄性不育被分为细胞质不育(质核互作不育)和细胞核不育两大类,并被广泛认可。
再后来的研究表明,核不育不仅仅只由隐性基因控制,植物界还普遍存在由显性核基因控制的雄性不育性,因而也可能有保持系(临时)和恢复系,在生产上也可以用来生产杂交种,如油菜、大白菜等,水稻也存在显性核不育现象,曾被较深入地研究。
20世纪70年代,湖北学者石明松又发现了后来定名的光周期敏感核不育,证明核不育水稻在特定温光条件下可以转换为可育,自交结实,实现一系两用,导致了“两系法”杂交水稻的诞生,同时丰富了植物雄性不育性的遗传理论,雄性不育性不仅仅受基因控制,有的还受基因与特定环境共同决定。可见,核不育也比较复杂。
雄性不育性除了细胞质不育和核不育两大类型以外,细胞质不育其花粉的育性又有孢子体(植株)基因型控制和配子体(花粉)基因型控制两大类型,即孢子体不育和配子体不育。核不育均为孢子体不育,即花粉的育性受植株基因型控制;理论上说,核不育不可能有配子体不育,因为如果有,就不可能产生不育株,遗传便终止了。
综上所述,水稻有应用价值的雄性不育性可分为细胞质不育(三系法)和细胞核不育(两系法),而细胞质不育又分为孢子体不育和配子体不育两类。我国育成并审定的水稻不育系分类和其主效基因归纳于表1。
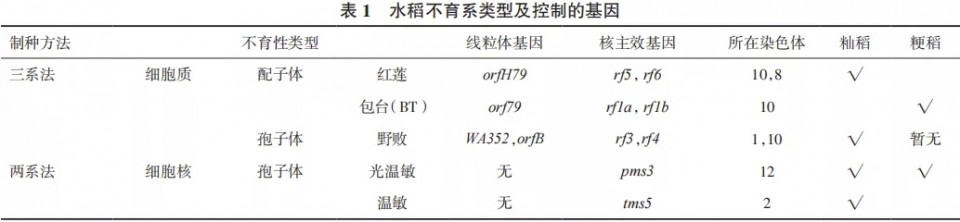
虽然我国不育系选育除了野败型、红莲型、BT(包台)型以外,还报道育成了矮败型、冈型、K型、马协型、D型、印尼水田谷型、滇型等60多种不育系和保持系,但从恢保关系、花粉败育时期及后期分子遗传学研究看来,均分别归类于野败、红莲、BT 3种类型。红莲、BT均属配子体不育,其恢保关系、败育时期均极为相似,分子遗传研究也表明,红莲型细胞质不育基因orfH79与已鉴定的BT型水稻不育基因orf79相比,orfH79上游(与atp6的间隔区)存在一个36bp的非同源序列,而编码区同源性高达97%。配子体雄性不育,在籼稻中主要是红莲型,在粳稻中主要是BT型;由于遗传机理的相近性,红莲型粳稻不育系、BT型籼稻不育系可能就显得意义不大,所以少有研究。对于光温敏核不育,既然pms3发源于粳稻,有无tms5也显得意义不大。但是,从表1可见,粳稻还没有孢子体不育的野败型不育系,这是粳稻不育系育种的一个空白,有待攻克。据了解,野败细胞质粳稻不育系已经育成,有商业价值的强优势恢复系也被创制出来,只是尚未公开报道而已。
★2.3 杂交种的选育
强优势杂交种的选育是杂交水稻育种至关重要的环节。从1973年广西的张先程、李丁民等同时最先发现IR24等强优势恢复系和袁隆平团队育成南优2号、颜龙安育成汕优2号等杂交稻品种以来,至2021年12月21日,国家水稻数据中心收录通过省级以上审定的杂交水稻品种7835个,其中籼型三系5062个,籼型两系2280个;粳型三系364个,粳型两系42个;“粳母籼父”型籼粳杂交稻83个,“籼母粳父”型籼粳杂交稻4个。
从数量上可以看出,亚种内品种间杂交稻品种选育跟不育系选育一样,在数量上也体现出籼稻远远多于粳稻、三系远远多于两系的局面。同时,还可以看出我国育种家还选育出了一些亚种间杂交种通过省级以上审定。其中,“粳母籼父”三系杂交种83个,“籼母粳父”三系杂交种2个、两系杂交种2个。这表明,三系法亚种间杂种优势利用主要集中于“粳母籼父”模式,三系“籼母粳父”模式才刚刚开始,两系“籼母粳父”模式似乎已经终结。
PART.3
水稻杂种优势利用路径剖析
★3.1 品种间杂种优势
杂种优势理论认为,杂种优势大小与双亲的遗传距离大小成正比。因此,人们推测,品种间的杂种优势<亚种间的杂种优势<种间的杂种优势。当然,随着双亲间遗传距离增加,其杂种优势利用的难度也越来越大,因此最容易的还是品种间杂种优势利用。我国水稻杂种优势利用率先突破的就是籼稻品种间的杂种优势利用,之后粳稻也获得突破。
就水稻而言,籼稻品种间的杂种优势明显大于粳稻品种间的杂种优势。这符合“杂种优势大小与双亲的遗传距离大小成正比”的规律,因为学术界多数人认为,籼稻由野生稻进化而来,粳稻是由早期的籼稻进化而来之后独立进化或驯化。
籼稻由于进化时间长,基因资源更为丰富,品种间的遗传距离更大。由于水稻品种间杂种优势利用已经接近50年,我国已经培育出7800多个杂交种,品种间杂种优势的挖掘可能达到了一个很高的高度,继续提高可能碰到了“天花板效应”,因此必须探索亚种间杂种优势利用。
★3.2 亚种间杂种优势
亚种间杂种优势大于品种间杂种优势,在理论上和实践中都是毫无疑问的。2005年宁波市农业科学院和宁波市种子公司用粳稻不育系甬粳2号A与籼型恢复系K4806(K2001/鄂恢)配制出甬优6号,并通过浙江省审定,标志着杂交水稻育种的新赛道——籼粳亚种间杂种优势利用正式开启。据不完全统计,2005年至2021年12月21日,国家水稻数据中心收录通过省级以上审定的“粳母籼父”杂交水稻品种83个,“籼母粳父”杂交稻品种4个。在2021年农业农村部公布的135个超级稻品种中,籼粳交杂交稻占据8席,这证明亚种间杂种优势利用不仅可能,而且非常有效,完全可能成为杂交水稻育种的新赛道。
★3.3 种间杂种优势
理论上说,种间杂种优势>亚种间杂种优势>品种间杂种优势。但是,亚种间就已经存在一定程度的生殖隔离,表现出杂交不亲和现象,种间就存在更加严重的杂交不亲和性。所以,种间杂种优势利用可能非常困难。
从我国杂交水稻育种的实践可以清楚地看出,中国杂交水稻育种的路径正在由品种间走向亚种间,以获得产量等性状更大的杂种优势。至于未来能否利用种间杂种优势,还有待科学和技术的进步、资源研究的突破,目前很难预测。既然动物中驴与马的杂交后代骡子就是种间杂种优势利用的实证,那么水稻的这种可能性在理论上也不能被完全排除。
PART.4
杂交水稻育种的新赛道
★4.1 籼粳交模式
杂交水稻育种自第一个杂交稻品种南优2号问世以来,已经接近50年,半个世纪的杂交水稻育种,基本上就是品种间杂种优势利用。50年的时间里,全国成百上千的研发机构、育种团队在拼搏,培育出审定品种7800多个,绝大多数局限于籼稻亚种中。时至今日,完全应该探索杂交水稻育种的新赛道。作者认为,无论是从理论上还是从浙江省多个育种团队的成功实践来看,籼粳亚种间杂交种的培育应该是杂交水稻育种的新赛道。
籼粳两个亚种间杂种优势的利用,因为不育系所属的亚种不同,有“粳母籼父”和“籼母粳父”两个模式。所谓“粳母籼父”模式,就是以粳稻不育系为母本、籼型恢复系为父本,生产F₁杂交种,供水稻生产使用。进而因不育系是“三系”还是“两系”,又有三系法“粳母籼父”模式和两系法“粳母籼父”模式两种情况。
三系法“粳母籼父”模式由宁波市农业科学院于2005年率先突破,选育出第一个粳籼交杂交种甬优6号并通过浙江省审定,之后便一发不可收拾,已育成至少66个亚种间杂交种且通过审定,并进行了较大面积应用,还有甬优12等6个品种被农业农村部认定为超级稻品种,占2021年全部有效超级稻品种数量的6/135。浙江省农业科学院、中国水稻研究所、嘉兴市农业科学院、金华市农业科学院等先后培育出浙优18(2012)、春优84(2013)、嘉禾优1号(2016)、长优2号(2017)等约20个粳籼交杂交种并通过审定。从目前审定的“粳母籼父”杂交种来看,主要是位于浙江省的国家、省、市研究机构和公司,其他省的育种团队似乎还没有审定品种。
两系法“粳母籼父”模式,由于粳稻两系不育系数量极少,仅区区8个,还包括最原始的农垦58S,“粳母籼父”模式的两系法粳籼交杂交种似乎为0,这也完全可以理解。
所谓“籼母粳父”模式,就是以籼稻不育系为母本、粳型恢复系为父本,生产F₁杂交种,供水稻生产使用。在理论上说,“籼母粳父”模式也有三系法“籼母粳父”模式和两系法“籼母粳父”模式两种情况。
三系法“籼母粳父”模式,似乎仅仅审定了2个品种,一个是中国水稻研究所育成的五优84(2015),一个是湖北省农业科学院育成的旱优79(2021)。其原因可能是恢复系创制涉及的基因太多,至少有广亲和基因S5n,恢复基因Rf3和Rf4或者Rf5和Rf6,还有不感光基因等,恢复系创制难度很大。
两系法“籼母粳父”模式亚种间杂交种选育,在理论上说,比三系法亚种间杂交种选育要容易得多,因为恢复系的创制不涉及恢复基因,仅涉及到广亲和基因,杂交种也不涉及到野生稻细胞质与粳稻细胞核基因组的远缘矛盾等问题。但由于粳稻感光性的问题,一般在9月才开始抽穗,而此时两系不育系已发生育性转换,难以找到合适的制种区域,所以尽管全国已培育出2000多个籼型两系不育系,但少有“籼母粳父”模式的两系法籼粳交杂交种。似乎只有信阳市农业科学院利用本身具有广亲和基因的培矮64S作母本,培育出两优培粳和信杂粳1号,并于2003年通过审定,或许由于制种的原因,这两个品种虽然在区域试验中增产幅度很大,但实际推广面积很小。
由此可见,两系法籼粳交杂种优势利用,无论是“粳母籼父”模式,还是“籼母粳父”模式,均由于不育系育性转换,实际中没有合适的制种区域,商业化应用似乎也没有可能。表2是我国籼粳交杂交稻育种的现状。
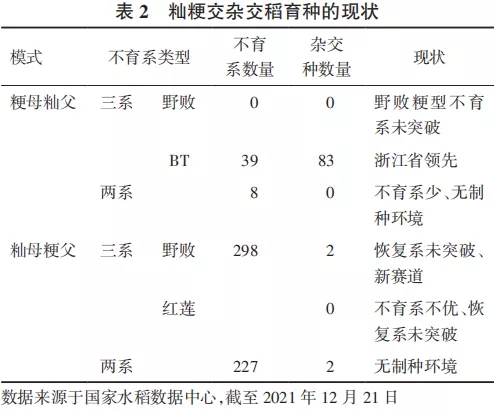
由上述分析可见,三系法“籼母粳父”亚种间杂种优势利用模式,成为杂交水稻育种新赛道中的新赛道。所以,有抱负的水稻育种团队,应该加大马力、踩足油门,在新赛道上狂奔,夺取杂交水稻育种的制高点。
★4.2 籼粳交不同模式比较
籼粳亚种间杂种优势利用有“籼母粳父”与“粳母籼父”两种模式,对这两种模式的异同进行比较(表3)。
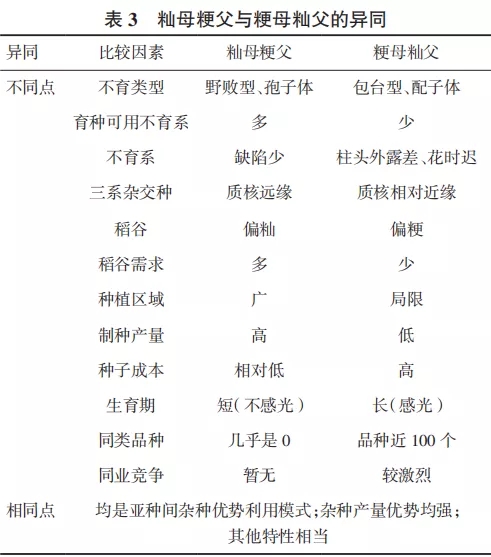
从表3可见,通过“籼母粳父”模式利用水稻亚种间杂种优势,有着许多优点和优势,值得探索和尝试。可以预见,鉴于籼稻不育系育种已有近50年的积累,通过省级审定的“三系”不育系和“两系”不育系分别多达298个和227个,是粳稻不育系数量的10倍之多,一旦其粳稻恢复系创制获得突破,籼型杂交稻50年的育种成果积淀就可能被深度开发和利用,水稻亚种间杂种优势利用便可以迅速上台阶。当然粳稻恢复系的创制其难度也是很大的。
(参考文献略)
本文选自《杂交水稻育种的新赛道》
作者:刘定富,应继锋
单位:武汉金玉良种科技有限公司;中国水稻研究所
刊于《中国种业》2022年第2期13-18页,转载请注明